
First Occurrence of Coffee Leaf Rust Caused by Hemileia vastatrix on Coffee in Saudi Arabia

1
Department of Arid Land Agricultural, College of Agricultural and Food Sciences, King Faisal University, Al-Ahsa 31982, Saudi Arabia
2
Pests and Plant Diseases Unit, College of Agricultural and Food Sciences, King Faisal University, Al-Ahsa 31982, Saudi Arabia
3
Vegetable Diseases Research Department, Plant Pathology Research Institute, Agricultural Research Center (ARC), Giza 12619, Egypt
*
Author to whom correspondence should be addressed.
Microbiol. Res. 2024, 15(1), 164-173; https://doi.org/10.3390/microbiolres15010011
Submission received: 11 December 2023
/
Revised: 28 December 2023
/
Accepted: 29 December 2023
/
Published: 8 January 2024
Abstract
:Coffee leaf rust (CLR) is a major disease of Arabica coffee caused by the biotrophic fungus Hemileia vastatrix. Jazan region in Saudi Arabia has long been one of the last coffee-producing regions in the world free of this disease. In August 2023, CLR was initially observed in coffee plantations located in Fyfa district one of the main coffee-producing mountains of Jazan region. The source of the infection is unknown, however CLR could have entered Jazan from the nearest coffee-producing locations that haven the pathogen. During a scheduled survey in August 2023, symptoms were observed including yellowish-orange lesions that frequently combined to form chlorotic lesions and exhibited the powdery appearance of yellow uredospores on the abaxial surface of leaves. The uredospores and teliospores were microscopically examined and their morphology matched the previously published description for H. vastatrix. The identity of H. vastatrix specimens was further confirmed based on PCR amplification and sequencing of ITS, sharing a 99–100% identity with previously published sequences, as belonging to H. vastatrix. The pathogenicity of H. vastatrix specimens was investigated on Coffee arabica plants under growth chamber conditions, and all were pathogenic relative to control, with 100% of disease incidence, therefore fulfilling Koch’s postulates. Based on our findings, this is the first documentation of H. vastatrix causing CLR in Saudi Arabia.
1. Introduction
One of the five largest families of flowering plants, the Rubiaceae comprises three subfamilies, 620 genera and 13,000 species, including coffee group species Arabica (Coffea arabica L.) and Coffea robusta L. Linden (C. robusta) (syn: Coffea canephora Pierr ex. A. Frohener (C. canephora)) are the two main types of coffee grown in the world [1]. Arabica accounts for 70% of coffee production, while Robusta accounts for 30% [2]. The domestication of Arabica coffee originated in Yemen, and its cultivation subsequently expanded to Asia, America, and other regions of Africa [3]. Arabica coffee has been grown in Yemen and Saudi Arabia for hundreds of years, on narrow valleys and terraced mountainsides at altitudes between 1200 and 1800 m [4]. In the Jazan, Asir, and Al-Baha locations of southwest Saudi Arabia, historic coffee-growing sites include trees that are over a century old [5]. According to Al-Asmari et al. [5], the Fyfa Development Authority (FDA), a governmental body, has reported that the approximate total count of trees is 78,000, with the majority (84%) being situated in Addayer district inside Jazan area. The yearly production of coffee beans is around 500 tons [4]. However, the number of trees is probably understated because the FDA runs a sizable nursery that provides local farmers with thousands of free seedlings every year (more than 40,000 coffee seedlings were given out in 2016 alone). Coffee in Saudi Arabia is primarily grown using organic cultivation methods without the use of synthetic pesticides, herbicides or fertilizers [6]. Farmers frequently incorporate organic goat dung with stone mulching [7]. In southwestern Saudi Arabia, coffee is a major socioeconomic factor and a source of revenue by the inhabitants of mountainous areas farmers. Research on coffee cultivation in the area was increased because of growing interest in coffee production [8]. Coffee farmers have several challenges in their coffee output across the tropics [9]. The production of coffee has been negatively affected by climate change either directly, as seen by a decrease in crop yield and quality, or indirectly, as shown by an increase in invasive pests and fungal, bacterial and viral diseases [10]. The three most common fungal diseases of coffee are Coffee Leaf Rust (CLR) caused by Hemileia vastatrix Berk. et Br. [11], Coffee Berry (CBD) caused by Colletotrichum coffeanum Noack (renamed C. kahawae) [12] and Coffee Wilt (CWD) caused by Fusarium xylarioides [13]. The coffee leaf rust that nearly wiped out coffee 150 years ago is one of the most virulent diseases and still causes issues for coffee plants across the world [14]. The causal agent was described as H. vastatrix [15]. As soon as it was first reported, the disease wiped out all coffee production areas in Ceylon (renamed “Sri Lanka”), which had terrible social and economic effects [16]. CLR has become one of the most well-known plant diseases in history since this sudden and terrible breakout. CLR can weaken the plant, causing branches to totally wither, and even cease the plant’s growth in severe cases. It can also cause damage that result in significant yield losses of up to 75% [17]. Despite recently published papers [18,19,20] which reported for the first time the association of certain fungi with diseases on coffee is Saudi Arabia, limited is known about the other threats to coffee production. In Jazan region, coffee farmers observed rust-like symptoms on several coffee trees with many trees heavily defoliated (personal interview). Upon the request of many farmers, a scheduled survey was conducted in August 2023 to assess the sanitary status of coffee plantations in Jazan region. Thus, this study used morphological and molecular approaches including phylogenetic analyses to identify the rust fungus causing disease on coffee trees in Jazan, Saudi Arabia.
2. Materials and Methods
2.1. Sampling and Morphological Identification
During August 2023, 15 commercial coffee plantations located in Fyfa district one of the main coffee-producing mountains of Jazan region were surveyed. Five trees were randomly monitored for each plantation by walking in a zig-zag pattern [21]. Ten leaves per tree were randomly collected from lateral branches exhibiting a substantial density of rust pustules and showing no visible signs of fungal diseases or insect infestation, were collected in plastic bags and brought to the laboratory of Pests and Plant Diseases Unit (PPDU) as soon as possible. In total, 750 leaves were collected and promptly examined using Olympus SZX10 stereomicroscope. Leaves exhibiting only rust colonies were chosen for subsequent investigation. After removing any impurities from the obtained leaves using a stereomicroscope, the spores were collected in an Eppendorf tube and kept at 4 °C for processing and further study [22]. Morphological examination of uredospores and teliospores was done after mounted in sterile distilled water using Leica DM2500 LED light microscope [22]. Furthermore, selected coffee leaves bearing H. vastatrix colonies were dried by pressing them between dry blotting papers and maintained at room temperature (25 ± 2 °C) according to Le et al. [23]. and deposited in the herbarium of Pests and Plant Diseases Unit (PPDUH), College of Agricultural and Food Sciences, King Faisal University, Saudi Arabia.
2.2. PCR Amplification and Sequencing
To confirm the identity of H. vastatrix-like fungi, DNA was extracted from uredospores using Dellaporta DNA isolation method [24]. The internal transcribed spacers (ITS) gene region, including the 5.8S nuclear ribosomal was amplified and sequenced using the primer pairs ITS1 and ITS4 [25]. PCR reactions were carried out in 25-µL reaction volume, which consisted of 10 µL PCR Master Mix (amaR OnePCR, GeneDirex, Inc., Las Vegas, NV, USA), 11 µL of ddH2O, 1.5 µL of each primer, and 1 µL of template DNA. The PCR amplification conditions were as follows: initial denaturation at 95 °C for 5 min, followed by 35 cycles of denaturation at 95 °C for 1 min, annealing at 54 °C for 30 s, and extension at 72 °C for 1 min, followed by a final extension at 72 °C for 10 min. The amplicons produced by PCR were sequenced in both directions in accordance with the manufacturer’s guidelines at Macrogen Company (Seoul, Republic of Korea). By utilizing the BLAST search (basic local alignment search tool) on GenBank, the most closely corresponding taxa were identified.
2.3. Phylogenetic Analysis
The generated sequences were edited, trimmed and aligned using MEGA v. 11.0.8 [26]. The taxonomic relationship of the H. vastatrix isolates was investigated through phylogenetic analysis of ITS gene region. The IQ-TREE multicore version 2.2.0 [27] was used to infer Maximum likelihood (ML) phylogenetic tree, based on 10,000 ultrafast bootstrap support replicates [28] and to calculate the best-fit evolution model according to BIC by ModelFinder [29]. Bayesian analysis was performed running MrBayes v3.2.6 on Cipres Science Gateway “www.phylo.org (accessed on 30 October 2023)”, on the combined, partitioned dataset with the substitution models, adapted by the previously ModelFinder calculation. A Bayesian analysis was conducted in duplicate using four Markov chain Monte Carlo (MCMC) chains, sampling every 1000 generations, and employing random trees for 10,000,000 generations. Bayesian phylogenetic trees and the ML tree (Figure 3) were observed using FigTree v. 1.4.4 (http://tree.bio.ed.ac.uk/software/figtree (accessed on 30 October 2023)). The generated sequences of the obtained specimens were deposited in GenBank (Table 1).
2.4. Pathogenicity Tests
In order to meet Koch’s postulates, the pathogenicity of H. vastatrix was investigated using the Keith et al. [30] method on apparently healthy 6-month-old C. arabica seedlings cv. khawlani. This cultivar is extensively cultivated in Jazan area and was found to be severely infected with H. vastatrix (personal interview). Uredospores were scraped from symptomatic coffee leaves and placed in a sterile Falcone tube with sterilized dH20. A droplet of Tween 80 was added to the spore suspension and vortexed to allow uredospore dispersion. Using a hemocytometer slide, the spore concentration was adjusted to 1 × 105 uredospores/mL in sterile water. A camel hair paintbrush was used to inoculate and spread the spore suspension to the abaxial side of coffee leaves [30]. Only sterile water was used to inoculate control plants. Plants were placed in a dark humid chamber at 22 ± 2 °C for 48 h before being switched to 12-h illumination under cool white fluorescent lights. Inoculated plants were monitored periodically for observing the developed pustules. The incidence of H. vastatrix was calculated after 30 days using the scale established by Julca-Otiniano et al. [31].
3. Results
3.1. Survey and Morphological Identification
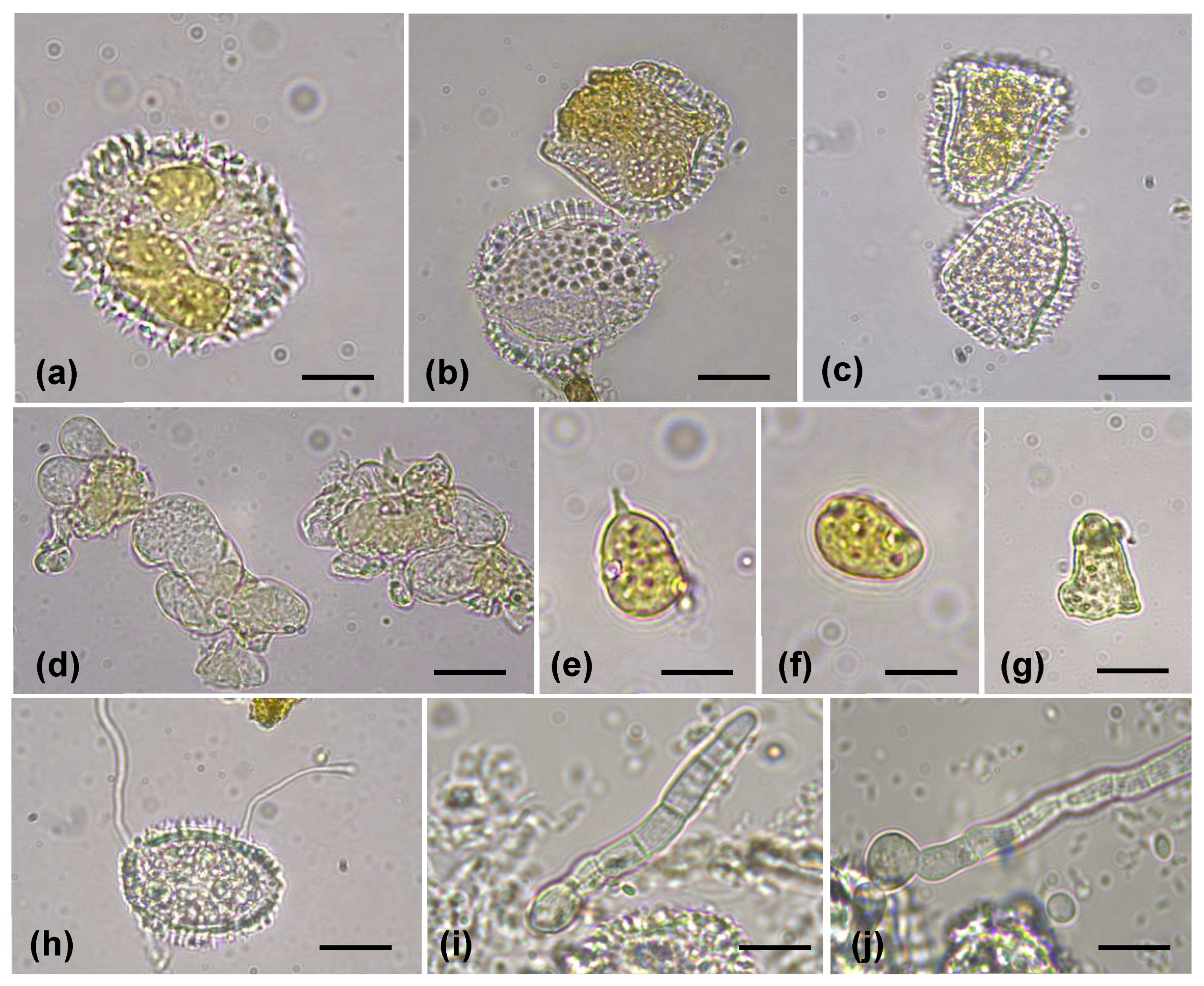
Our investigation at 15 coffee plantations in the mountainous area of Fyfa district revealed that coffee rust disease incidence in this region was high and recorded a 100%. Across all 15 surveyed coffee plantations, the incidence of CLR in the lower canopy was found to have significantly higher incidence relative to mid-canopy branches. Moderate defoliation was observed in certain trees in some uncleaned farms. The monitored coffee trees displayed classic symptoms of round, yellowish-orange lesions with varying degree of severity, and on the abaxial surface exhibited the powdery appearance of yellow uredospores (Figure 1a,b). Close focus on the upper surface showing tissue necrosis in the center of the lesion (Figure 1c) and abaxial surface showing the powdery appearance of uredospores (Figure 1d). Microscopical examination revealed that uredospores were ovoid to reniform-shaped, convex on the dorsal side with a smooth hyaline to pale yellow-orange wall on the ventral side, finely to coarsely echinulate, measuring (25) 26.7–35 (35.4) × (16.7) 18.5–23.7 (24.6) µm (av. = 29.5 ± 2.70 × 21.6 ± 2.13 μm) (Figure 2a–c). Teliospores were spherical to angular-globose and mostly irregular, with a smooth pale-yellow wall, sometimes guttulate, measuring (10.52) 10.78–21.89 (24.15) × (8.04) 8.47–15.99 (17.13) µm (av. = 17.24 ± 4.03 × 11.96 ± 2.23 μm) (Figure 2d–g).
3.2. Phylogenetic Analysis
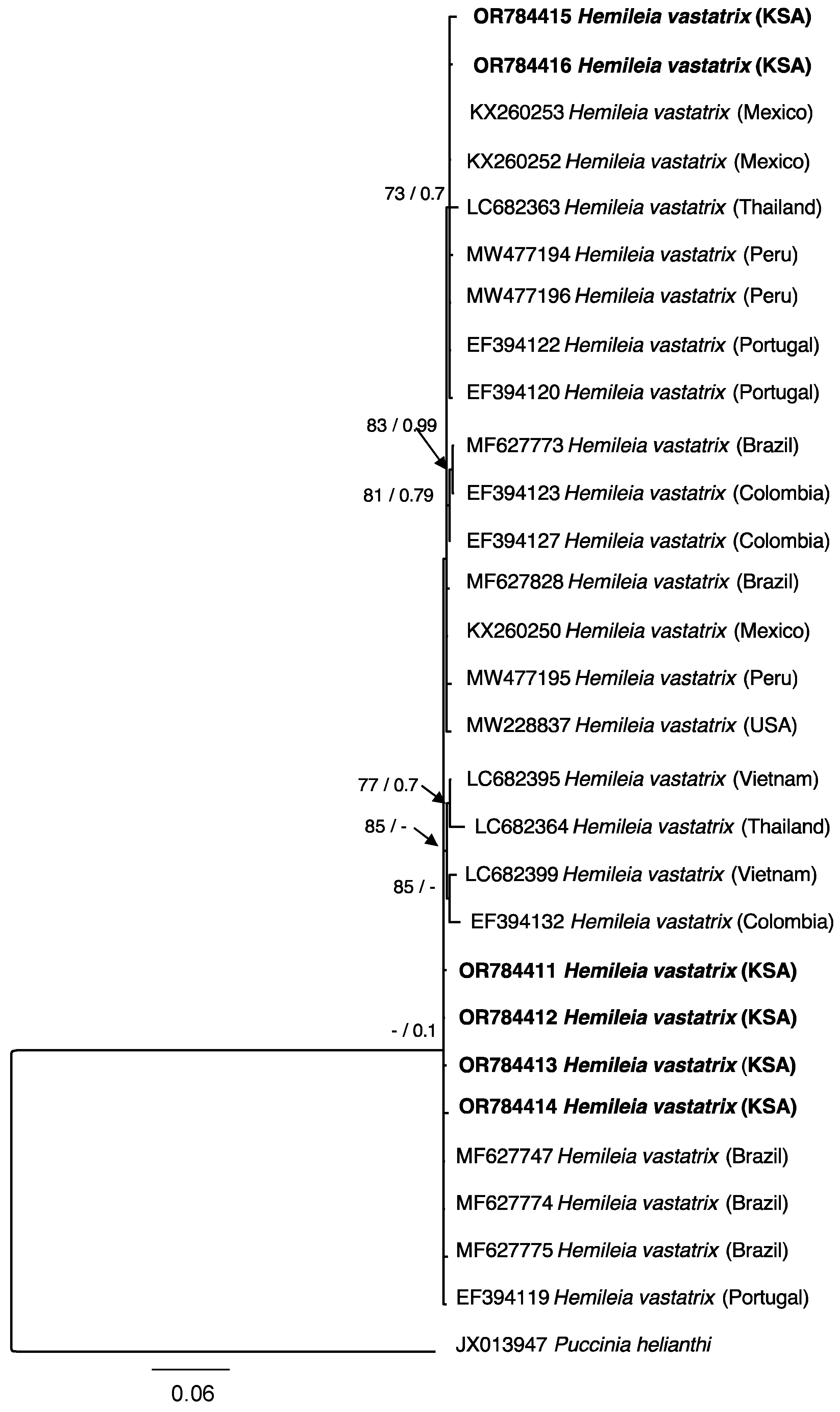
The identity of 17 H. vastatrix specimens collected in this study was further confirmed based on ITS sequencing, with generated sequences sharing 99–100% identity with publicly available sequences identified as H. vastatrix. The results of a phylogenetic analysis based on the ITS for representative six specimens and sequences retrieved from GenBank (Table 1) is illustrated in Figure 3. Among the six specimens, two of them were located far apart from the others within specimens from Mexico (Accession No: KX260252 and KX260253), Thailand (LC682363), Peru (MW477194 and MW477196), and Portugal (EF394122 and EF394120) (Figure 3). The remaining four specimens were grouped in a clade highly supported with posterior probability value BPP (1.0) within four specimens (Accession No: MF627747, MF627774 and MF627775) from Brazil and single specimen (EF394119) from Portugal.
3.3. Pathogenicity Tests
The pathogenicity of the collected uredospores was evaluated on 6-month-old coffee seedlings. Rust symptoms appeared first after 25 days as chlorotic halos on the upper side of leaves with small pustules contained orange uredospores on the abaxial surface [15]. By the day 30, the lesions exhibited an increase in size, subsequently leading to necrosis in the central region of many leaf lesions [32]. There was no variation in the final incidence of rust among all the inoculated plants, and the incidence was 100%. There were no premature defoliated leaves observed, and leaf abscission was recognized 65 days later [30]. The identity of uredospores recovered from the observed lesions was confirmed as H. vastatrix by microscopic examination, fulfilling Koch’s postulates.
4. Discussion
Coffee farmers across the different regions of southwest Saudi Arabia have several challenges in their agricultural output. The production of coffee has been impacted by climate change either directly, as seen by a decrease in crop yield and quality, or indirectly, as shown by an increase in invasive pests and diseases [19,20]. The results presented here provide new information about the identification and documentation of the presence of H. vastatrix on coffee in Saudi Arabia. We found no significant difference in CLR incidence among coffee plantations, suggesting that environmental factors are generally not limiting to CLR survival and germination on commercial farms in Jazan. We also found that low-canopy branches had a much greater CLR incidence than mid-canopy branches, which was reflected in the pattern of defoliation, with lower branches losing leaves first, followed by mid-canopy branches as the infection progressed. The initial defoliation of the lower branches is most likely caused by self-shading within trees, which can create optimal circumstances for CLR survival and germination by restricting sun exposure and prolonging the period of leaf wetness [33]. The typical symptoms of CLR were clearly observed and coincided those described in many studies [18,34]. The morphology of the examined uredospores and teliospores matched the previous descriptions for H. vastatrix Ward [35,36,37]. Both uredospores and teliospores were produced together in the same pustule. However, the incidence of teliospores was scarce and observed in very low frequency compared to the uredospores. This might be hampered by the environmental conditions during the survey period. This finding is supported by those of Fernandes et al. [36]. who demonstrated that occurrence of teliospores was of 20% in July, 15% in August, and 5% in September. The internal transcribed spacer (ITS) region of the ribosomal DNA (rDNA) has been extensively recognized as a primary barcode marker for fungi [38]. In this study, the ITS sequence data was sufficient to help in identification the six specimens of H. vastatrix. Furthermore, the utilization of rDNA-ITS as a reliable marker has been emphasized for distinguishing not only at the taxonomic level of species but also among individuals within a population [39]. The findings derived from the phylogenetic analysis indicate that the six specimens of the H. vastatrix population exhibit a lack of structure, indicating a homogenous population with no discernible differentiation based on geographic origin. The undifferentiated H. vastatrix population could have evolved from the movement of spores between the two areas transported by people, seed, and/or infected plants [40]. These findings are consistent with those obtained in Brazil by Cabral et al. [41], who found no evidence of H. vastatrix populations being structured based on origin, host, or physiological race. However, the genetic diversity was lower than what was revealed in the current study. Using 91 fungal isolates, Maia et al. [42] achieved results comparable to the current investigation; they argued that H. vastatrix acts as a huge undifferentiated population with significant genotypic diversity that is not structured based on its geographic origin and host. As a result, the fungus’s spread over extensive distances may explain the absence of structure in the six H. vastatrix populations by region. The lack of population structuring in H. vastatrix, with respect to geographic origin, may be attributed to a substantial gene flow facilitated by the transportation of infected plant materials from the nearby Yemen Mountains [43]. It is suggested that wind had a significant role in the dissemination of pathogens across large distances, and has been proposed as the primary factor behind the introduction of coffee leaf rust to coffee farms in Saudi Arabia [44]. However, we cannot ascertain that the H. vastatrix population in Saudi Arabia spread from Yemen Mountains. The origin of CLR is believed to have been in eastern Africa. It was reported in 1869 in Ceylon (now Sri Lanka) and by the 1920s, it had spread to most parts of Africa and Asia [12,45]. In Kenya, CLR was first reported in 1912 [46]. The emergence of CLR in Ethiopia was formally documented in 1934, albeit without causing the epidemics that have been documented in other regions [47]. Upon further consideration, it is plausible that H. vastatrix may have been carried by the wind to Saudi Arabia from neighboring African countries, given the proximity of these countries to Yemen and Saudi Arabia. A notable instance of this kind of dispersal mechanism is the yellow rust epidemics caused by a virulent strain of Puccinia striiformis, known as Yr9, which originated in eastern Africa and gradually spread to South Asia via the Middle East and West Asia over a period of approximately ten years [48]. These findings suggest that H. vastatrix may have been migrated by the wind to production areas in Jazan region and surrounding areas.
Coffee has been cultivated in the south-western region of Saudi Arabia for several centuries, mostly through seed propagation (personal interview). Consequently, it is likely that the genotypes referred to local producers in this area are, in fact, populations consisting of numerous accessions, exhibiting varying degrees of genetic diversity [49]. Therefore, there is a significant change in the frequency and distribution of the resistant local coffee genotypes to H. vastatrix [49]. The latter is due to the coffee plantation replacement plan is likely to have been caused by the Saudi government to control and replace invasive Khat or qat trees (Catha edulis) with coffee trees. The population structure of H. vastatrix in Saudi Arabia has not yet been investigated and this study is the first to report the occurrence of this pathogen on coffee in Saudi Arabia. However, the diversity of the H. vastatrix population may be subject to selection pressure from the resistance genes of the cultivated coffee cultivars, which could lead to the introduction of new pathogen variations. As a result, there is currently a good chance that new pathogen variations will appear [50]. Therefore, it is crucial to investigate the genetic diversity and population structure of plant pathogens in order to monitor pathogen evolution, create disease control measures, and understand fungal epidemiology [41]. Due to the constraint imposed by the limited number of samples, our future research endeavors will focus on conducting population genomic studies on H. vastatrix in Saudi Arabia. The objective of these studies will be to gain further insights on the genetic structure, population demographics, origins, and migration patterns of H. vastatrix within the Saudi Arabian context.
Author Contributions
Conceptualization, A.M.I. and K.A.; methodology, A.M.I.; software, A.M.I. and K.A.; writing—original draft preparation, A.M.I. and K.A.; writing—review and editing, K.A. and A.M.I. All authors have read and agreed to the published version of the manuscript.
Funding
This work is financed by the Deanship of Scientific Research, Vice Presidency for Graduate Studies and Scientific Research, King Faisal University, Saudi Arabia, through grant number (GRANT5425).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data related to this study is mentioned in the manuscript.
Acknowledgments
Authors extend their gratefulness to the Deanship of Scientific Research, Vice Presidency for Graduate Studies and Scientific Research, King Faisal University, Saudi Arabia, for supporting this research for work through grant number (GRANT5425). We would like to acknowledge the technical staff Eng. Mustafa I. Almaghasla for his assistance in the molecular analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Govaerts, R.; Ruhsam, M.; Andersson, L.; Robbrecht, E.; Bridson, D.M.; Davis, A.P.; Schanzer, I.; Sonké, B. World Checklist of Rubiaceae. Royal Botanic Gardens, Kew. 2011. Available online: http://www.kew.org/wcsp/rubiaceae (accessed on 22 September 2023).
- Sahachairungrueng, W.; Meechan, C.; Veerachat, N.; Thompson, A.K.; Teerachaichayut, S. Assessing the Levels of Robusta and Arabica in Roasted Ground Coffee Using NIR Hyperspectral Imaging and FTIR Spectroscopy. Foods 2022, 11, 3122. [Google Scholar] [CrossRef] [PubMed]
- Montagnon, C.; Rossi, V.; Guercio, C.; Sheibani, F. Vernacular Names and Genetics of Cultivated Coffee (Coffea arabica) in Yemen. Agronomy 2022, 12, 1970. [Google Scholar] [CrossRef]
- Tounekti, T.; Mosbah, M.; Al-Turki, T.; Khemira, H. Genetic diversity analysis of Coffee (Coffea arabica L.) germplasm accessions growing in the Southwestern Saudi Arabia using quantitative traits. Nat. Resour. 2017, 8, 321–336. [Google Scholar]
- Al-Asmari, K.; Abu Zeid, I.; Al-Attar, A. Coffee Arabica in Saudi Arabia: An Overview. Int. J. Pharm. Phytopharm. Res. 2020, 10, 71–78. [Google Scholar]
- Lewin, B.; Giovannucci, D.; Varangis, P. Coffee Markets: New Paradigms in Global Supply and Demand; (SSRN Scholarly Paper No. ID 996111); Social Science Research Network: Rochester, NY, USA, 2004. [Google Scholar]
- Al-Turki, T.A. An initiative in exploration and management of plant genetic diversity in Saudi Arabia. In Managing Plant Genetic Diversity, Proceedings of the International Conference, Kuala Lumpur, Malaysia, 12–16 June 2000; CABI Publishing: Wallingford, UK, 2002; pp. 339–349. [Google Scholar]
- Tounekti, T.; Mahdhi, M.; Al-Turki, T.A.; Khemira, H. Water relations and photo-protection mechanisms during drought stress in four coffee (Coffea arabica) cultivars from southwestern Saudi Arabia. S. Afr. J. Bot. 2018, 117, 17–25. [Google Scholar] [CrossRef]
- Harvey, C.A.; Rakotobe, Z.L.; Rao, N.S.; Dave, R.; Razafimahatratra, H.; Rabarijohn, R.H.; Rajaofara, H.; MacKinnon, J.L. Extreme vulnerability of smallholder farmers to agricultural risks and climate change in Madagascar. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130089. [Google Scholar] [CrossRef] [PubMed]
- Pham, Y.; Reardon-Smith, K.; Mushtaq, S.; Cockfield, G. The impact of climate change and variability on coffee production: A systematic review. Clim. Chang. 2019, 156, 609–630. [Google Scholar] [CrossRef]
- Waller, J.M. Coffee rust-epidemiology and control. Crop Prot. 1982, 1, 385–404. [Google Scholar] [CrossRef]
- Waller, J.M.; Bridge, P.D.; Black, R.; Hakiza, G. Characterization of the coffee berry disease pathogen, Colletotrichum kahawae sp. nov. Mycol. Res. 1993, 97, 989–994. [Google Scholar] [CrossRef]
- Adugna, G.; Million, A.; Hindorf, H.; Zeru, A.; Teferi, D.; Jefuka, C. Coffee wilt Disease in Ethiopia; CAB International: London, UK, 2010. [Google Scholar]
- Talhinhas, P.; Batista, D.; Diniz, I.; Vieira, A.; Silva, D.N.; Loureiro, A.; Tavares, S.; Pereira, A.P.; Azinheira, H.G.; Guerra-Guimarães, L.; et al. The coffee leaf rust pathogen Hemileia vastatrix: One and a half centuries around the tropics. Mol. Plant Pathol. 2017, 18, 1039–1051. [Google Scholar] [CrossRef]
- Berkeley, M.J.; Broome, C.E. Hemileia vastatrix. Gard. Chron. 1869, 6, 1157. [Google Scholar]
- Morris, D. Note on the structure and habit of Hemileia vastatrix, the coffee-leaf disease of Ceylon and Southern India 1880. J. Linn. Soc. Bot. 1880, 17, 512–517. [Google Scholar] [CrossRef]
- Gichuru, E.; Alwora, G.; Gimase, J.; Kathurima, C. Coffee Leaf Rust (Hemileia vastatrix) in Kenya—A Review. Agronomy 2021, 11, 2590. [Google Scholar] [CrossRef]
- Alhudaib, K.; Ismail, A.M.; Magistà, D. Multi-Locus Phylogenetic Analysis Revealed the Association of Six Colletotrichum Species with Anthracnose Disease of Coffee (Coffea arabica L.) in Saudi Arabia. J. Fungi 2023, 9, 705. [Google Scholar] [CrossRef] [PubMed]
- AL-Faifi, Z.; Alsolami, W.; Abada, E.; Khemira, H.; Almalki, G.; Modafer, Y. Fusarium oxysporum and Colletotrichum musae Associated with Wilt Disease of Coffea arabica in Coffee Gardens in Saudi Arabia. Can. J. Infect. Dis. Med. Microbiol. 2022, 2022, 3050495. [Google Scholar] [CrossRef] [PubMed]
- El-Komy, M.H.; Alsubaie, M.; Ibrahim, Y.E.; Sharafaddin, A.H.; Al-Saleh, M.A. First Report of Colletotrichum gloeosporioides and Colletotrichum siamense Causing Anthracnose Disease on Coffee Trees in Saudi Arabia. Plant Dis. 2023, 107, 3297. [Google Scholar] [CrossRef] [PubMed]
- Aristizábal, L.F.; Johnson, M.A. Monitoring Coffee Leaf Rust (Hemileia vastatrix) on Commercial Coffee Farms in Hawaii: Early Insights from the First Year of Disease Incursion. Agronomy 2022, 12, 1134. [Google Scholar] [CrossRef]
- Gamarra Gamarra, D.P.; Torres Suarez, G.; Villar Quiñonez, C.M.; McTaggart, A.R.; Carrasco Lozano, E.C. Phylogenetic relationship of coffee leaf rust in the central jungle of Peru. Acta Agronómica 2021, 70, 155–162. [Google Scholar] [CrossRef]
- Le, C.T.M.; Okane, I.; Ono, Y.; Tsuda, Y.; Yamaoka, Y. Incidence of Coffee Leaf Rust in Vietnam, Possible Original Sources and Subsequent Pathways of Migration. Front. Plant Sci. 2022, 13, 872877. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A Plant DNA Minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protoc. Guid. Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Keith, L.M.; Sugiyama, L.S.; Brill, E.; Adams, B.L.; Fukada, M.; Hoffman, K.M.; Ocenar, J.; Kawabata, A.; Kong, A.T.; McKemy, J.M.; et al. First report of coffee leaf rust caused by Hemileia vastatrix on coffee (Coffea arabica) in Hawaii. Plant Dis. 2022, 106, 761. [Google Scholar] [CrossRef] [PubMed]
- Julca-Otiniano, A.; Borjas-Ventura, R.; Alvarado-Huamán, L.; Julca-Vera, N.; Bello-Amez, S.; Castro-Cepero, V. Relationship between the incidence and severity of coffee rust (Hemileia vastatrix) in San Ramón, Chanchamayo, Peru. J. Sci. Res. 2019, 4, 1–9. [Google Scholar]
- Várzea, V.; Pereira, A.P.; Silva, M.d.C.L.d. Screening for Resistance to Coffee Leaf Rust. In Mutation Breeding in Coffee with Special Reference to Leaf Rust; Ingelbrecht, I.L., Silva, M.d.C.L.d., Jankowicz-Cieslak, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2023. [Google Scholar] [CrossRef]
- Lopez-Bravo, D.F.; de Virginio-Filho, E.M.; Avelino, J. Shade is conducive to coffee leaf rust as compared to full sun exposure under standardized fruit load conditions. Crop Prot. 2012, 38, 21–29. [Google Scholar] [CrossRef]
- Bekele, K.B.; Senbeta, G.A.; Garedew, W.; Caixeta, E.T.; Ramírez-Camejo, L.A.; Aime, M.C. Genetic diversity and population structure of Hemileia vastatrix from Ethiopian Arabica coffee. Arch. Phytopathol. Plant Prot. 2022, 55, 1483–1503. [Google Scholar] [CrossRef]
- Ward, H.M. On the morphology of Hemileia vastatrix Berk. and Br. (the fungus of the Coffee disease of Ceylon). Q. J. Microsc. Sci. 1889, 22, 1–11. [Google Scholar] [CrossRef]
- Fernandes, R.D.C.; Evans, H.C.; Barreto, R.W. Confirmation of the occurrence of teliospores of Hemileia vastatrix in Brazil with observations on their mode of germination. Trop. Plant Pathol. 2009, 34, 108–113. [Google Scholar] [CrossRef]
- Coutinho, T.A.; Rijkenberg, F.H.J.; Van Asch, M.A.J. Teliospores of Hemileia vastatrix. Mycol. Res. 1995, 99, 932–934. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Liu, B.; Li, L.; Huang, X.; Liang, Y.; Zheng, J.; Wang, Q.; Li, R.; He, C.; Yi, K. The rDNA-ITS sequences analysis and phylogenetic relationships of Hemileia vastatrix. Chin. J. Trop. Crops 2018, 39, 2250–2258. [Google Scholar]
- Quispe-Apaza, C.S.; Mansilla-Samaniego, R.C.; López-Bonilla, C.F.; Espejo-Joya, R.; Villanueva-Caceda, J.; Monzón, C. Genetic diversity of Hemileia vastatrix of two coffee producing areas in Peru. Rev. Mex. Fitopatol. 2017, 35, 418–436. [Google Scholar]
- Cabral, P.G.C.; Maciel-Zambolim, E.; Oliveira, S.A.S.; Caixeta, E.T.; Zambolim, L. Genetic diversity and structure of Hemileia vastatrix populations on Coffea spp. Plant Pathol. 2016, 65, 196–204. [Google Scholar] [CrossRef]
- Maia, T.A.; Maciel-Zambolim, E.; Caixeta, E.T.; Mizubuti, E.S.G.; Zambolim, L. The population structure of Hemileia vastatrix in Brazil inferred from AFLP. Australas. Plant Pathol. 2013, 42, 533–542. [Google Scholar] [CrossRef]
- Schieber, E. Economic impact of coffee rust in Latin America. Annu. Rev. Phytopathol. 1972, 10, 491–510. [Google Scholar] [CrossRef]
- Bowden, J.; Gregory, P.H.; Johnson, C.G. Possible wind transport of coffee leaf rust across the Atlantic Ocean. Nature 1971, 224, 500–501. [Google Scholar] [CrossRef]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The still underestimated problem of fungal diseases worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef]
- Silva, M.D.C.; Várzea, V.; Guerra-Guimarães, L.; Gil Azinheira, H.; Fernandez, D.; Petitot, A.-S.; Bertrand, B.; Lashermes, P.; Nicole, M. Coffee resistance to the main diseases: Leaf rust and coffee berry disease. Braz. J. Plant Physiol. 2006, 18, 119–147. [Google Scholar] [CrossRef]
- Sylvain, P.G. Some observations on Coffea arabica L. in Ethiopia. Turrialba 1958, 5, 37–54. [Google Scholar]
- Singh, R.P.; Hodson, D.P.; Jin, Y.; Huerta-Espino, J.; Kinyua, M.G.; Wanyera, R.; Njau, P.; Ward, R.W. Current status, likely migration and strategies to mitigate the threat to wheat production from race Ug99 (TTKS) of stem rust pathogen. CABI Rev. 2006, 2006, 1–13. [Google Scholar]
- Al-Ghamedi, K.; Alaraidh, I.; Afzal, M.; Mahdhi, M.; Al-Faifi, Z.; Oteef, M.D.Y.; Tounekti, T.; Alghamdi, S.S.; Khemira, H. Assessment of Genetic Diversity of Local Coffee Populations in Southwestern Saudi Arabia Using SRAP Markers. Agronomy 2023, 13, 302. [Google Scholar] [CrossRef]
- Quispe-Apaza, C.; Mansilla-Samaniego, R.; Espejo-Joya, R.; Bernacchia, G.; Yabar-Larios, M.; López-Bonilla, C. Spatial and temporal genetic diversity and population structure of Hemileia vastatrix from Peruvian Coffee Plantations. Plant Pathol. J. 2021, 37, 280. [Google Scholar] [CrossRef]
Figure 1.
(a) Symptoms of coffee leaf rust (CLR) on the upper leaf surface as round, yellowish-orange lesions; (b) symptoms on the lower leaf surface as powdery orange lesions; (c) close focus on the upper lesions with tissue necrosis occurs; (d) close focus on the abaxial surface showing the powdery appearance of uredospores.
Figure 1.
(a) Symptoms of coffee leaf rust (CLR) on the upper leaf surface as round, yellowish-orange lesions; (b) symptoms on the lower leaf surface as powdery orange lesions; (c) close focus on the upper lesions with tissue necrosis occurs; (d) close focus on the abaxial surface showing the powdery appearance of uredospores.

Figure 2.
(a–c) Uredospores of H. vastatrix; (d) Teliospores aggregated and flattended among uredospores; (e–g) Teliospores with and without papillae; germination of uredospores (h) and teliospores (i,j)—Scale bars = 10 µm.
Figure 2.
(a–c) Uredospores of H. vastatrix; (d) Teliospores aggregated and flattended among uredospores; (e–g) Teliospores with and without papillae; germination of uredospores (h) and teliospores (i,j)—Scale bars = 10 µm.

Figure 3.
Maximum likelihood tree obtained through heuristic searches of the ITS gene region of the H. vastatrix. Values of Bayesian posterior probability (BPP) and support values of Bootstrap (BS) (1000 replicates) are provided at the nodes. Branches that are unsupported with BS or BPP are denoted by –. Puccinia helianthi is treated as an outgroup. The sequences obtained in the current study are indicated in black boldface.
Figure 3.
Maximum likelihood tree obtained through heuristic searches of the ITS gene region of the H. vastatrix. Values of Bayesian posterior probability (BPP) and support values of Bootstrap (BS) (1000 replicates) are provided at the nodes. Branches that are unsupported with BS or BPP are denoted by –. Puccinia helianthi is treated as an outgroup. The sequences obtained in the current study are indicated in black boldface.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Sequences retrieved from GenBank for phylogenetic analysis and those obtained in this study.
Table 1.
Sequences retrieved from GenBank for phylogenetic analysis and those obtained in this study.
| Species Identity | Isolate No. | Host | Country of Origin | Accession Numbers |
|---|---|---|---|---|
| ITS | ||||
| H. vastatrix | PPDU1023 | C. arabica | Saudi Arabia | OR784411 |
| H. vastatrix | PPDU1123 | C. arabica | Saudi Arabia | OR784412 |
| H. vastatrix | PPDU1223 | C. arabica | Saudi Arabia | OR784413 |
| H. vastatrix | PPDU1323 | C. arabica | Saudi Arabia | OR784414 |
| H. vastatrix | PPDU1423 | C. arabica | Saudi Arabia | OR784415 |
| H. vastatrix | PPDU1523 | C. arabica | Saudi Arabia | OR784416 |
| H. vastatrix | Mexico_1 | Coffea arabica L., var. Caturra Rojo | Mexico | KX260253 |
| H. vastatrix | HVV_17.4_2018 | C. arabica | Peru | MW477196 |
| H. vastatrix | Mexico_2 | C. arabica var. Pluma Hidalgo | Mexico | KX260252 |
| H. vastatrix | CIFC_1 | C. arabica var. Caturra | Portugal | EF394122 |
| H. vastatrix | TL_18TH13 | Coffea sp. | Thailand | LC682363 |
| H. vastatrix | HVV_17.2_2018 | C. arabica | Peru | MW477194 |
| H. vastatrix | CIFC_2 | C. arabica var. Caturra | Portugal | EF394120 |
| H. vastatrix | Brazil_1 | C. arabica | Brazil | MF627828 |
| H. vastatrix | HVV_17.3_2018 | C. arabica | Peru | MW477195 |
| H. vastatrix | Haiku | C. arabica | USA | MW228837 |
| H. vastatrix | CMPH-165 | Coffea canephora var. Robusta | Mexico | KX260250 |
| H. vastatrix | Piranga_2 | C. arabica | Brazil | MF627773 |
| H. vastatrix | Colombia_10 | C. arabica var. Caturra | Colombia | EF394123 |
| H. vastatrix | Colombia_6 | C. arabica var. Colombia | Colombia | EF394127 |
| H. vastatrix | VNCR_S1_14 | Coffea sp. | Vietnam | LC682399 |
| H. vastatrix | Colombia_1 | C. arabica var. Caturra | Colombia | EF394132 |
| H. vastatrix | TL_19TH46 | Coffea sp. | Thailand | LC682364 |
| H. vastatrix | VNCR_S1_2 | Coffea sp. | Vietnam | LC682395 |
| H. vastatrix | CIFC_3 | C. arabica var. Caturra | Portugal | EF394119 |
| H. vastatrix | B1_1 | C. arabica | Brazil | MF627747 |
| H. vastatrix | Piranga_3 | C. arabica | Brazil | MF627774 |
| H. vastatrix | Piranga_4 | C. arabica | Brazil | MF627775 |
| Puccinia helianthi | NM-1 | Helianthus annuus L. (Sunflower) | China | JX013947 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alhudaib, K.; Ismail, A.M. First Occurrence of Coffee Leaf Rust Caused by Hemileia vastatrix on Coffee in Saudi Arabia. Microbiol. Res. 2024, 15, 164-173. https://doi.org/10.3390/microbiolres15010011
AMA Style
Alhudaib K, Ismail AM. First Occurrence of Coffee Leaf Rust Caused by Hemileia vastatrix on Coffee in Saudi Arabia. Microbiology Research. 2024; 15(1):164-173. https://doi.org/10.3390/microbiolres15010011
Chicago/Turabian StyleAlhudaib, Khalid, and Ahmed Mahmoud Ismail. 2024. "First Occurrence of Coffee Leaf Rust Caused by Hemileia vastatrix on Coffee in Saudi Arabia" Microbiology Research 15, no. 1: 164-173. https://doi.org/10.3390/microbiolres15010011